- Главная
- Авиация и космонавтика
- Административное право
- Акционерное право
- Английский
- Антикризисный менеджмент
- Биографии
- Автомобильное хозяйство
- Автотранспорт
- Культура и искусство
- Маркетинг
- Международное публичное право
- Международное частное право
- Международные отношения
- Менеджмент
- Металлургия
- Муниципальное право
- Налогообложение
- Оккультизм и уфология
- Педагогика
- Политология
- Право
- Предпринимательство
- Психология
- Радиоэлектроника
- Риторика
- Социология
- Статистика
- Страхование
- Строительство
- Схемотехника
- Таможенная система
- Теория государства и права
- Теория организации
- Теплотехника
- Технологии
- Товароведение
- Транспорт
- Трудовое право
- Туризм
- Уголовное право и процесс
- Управление
- Сочинения по литературе и русскому языку
- Другое
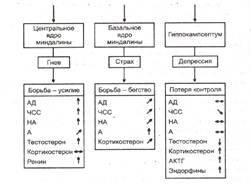
Курсовая работа: Влияние эмоций на организмКурсовая работа: Влияние эмоций на организмСодержание Введение 1. Нейроанатомия эмоций 2. Теория Хенри Список используемой литературы Введение Обычно эмоцию определяют как особый вид психических процессов, которые выражают переживание человеком его отношения к окружающему миру и самому себе – эта оценка влияет на последующие действия. Особенность эмоций состоит в том, что они в зависимости от потребностей субъекта непосредственно оценивают значимость действующих на индивид объектов и ситуаций. Эмоции выполняют функции связи между действительностью и потребностями. При этом также очевидно, что различные эмоции оказывают огромное влияние на организм в целом, т.к. даже при незначительных проявлениях чувств и эмоций в человеческом организме происходят различные внешние (мимика, пантомимика (поза), двигательные реакции, голос) и внутренние изменения (вегетативных показателей (частота сердечных сокращений, артериальное давление, частота дыхания и т.д.), которые по обратной связи отражаются на общем физическом состоянии и настроение. Итог, действия человека небеспристрастны, поэтому эмоция как субъективное переживание присутствует в составе каждой деятельности, каждого рефлекса, но каким же образом происходит воздействие эмоций на организм? На этот вопрос мы сможем ответить, если изучим представленные концепции и нейроанатомию эмоций. 1. Нейроанатомия эмоций Наиболее убедительные данные о нейроанатомии эмоций получены в отношении тех мозговых структур, которые определяют знак эмоций. Опыты на животных с электрической стимуляцией и разрушением мозга, а затем и с самостимуляцией мозговых структур показали существование двух типов центров, возбуждение которых вызывает эмоции с противоположным знаком (приятные и неприятные). Впервые об этом заговорили в 1954 г. на Международном физиологическом конгрессе в Монреале (Канада). П. Милнер и Дж. Олдс продемонстрировали эффект самораздражения у крыс. Вживив электроды в медиальный переднемозговой пучок в области латерального гипоталамуса, они поместили крысу в ящик, где находился рычаг, при нажиме на который в мозг крысы посылался удар тока. Перемещаясь в клетке, крыса случайно задевала рычаг, производя таким образом самораздражение этого участка мозга. Со временем крыса все больше времени проводила у рычага и все чаще производила нажимы на него, создавая постоянное самораздражение, когда частота нажима на рычаг достигала до 7 тысяч за один час. Стимуляция зон самораздражения вызывает положительные эмоциональные реакции и служит более сильным подкреплением, чем даже пищевое. На основе этого подкрепления легко вырабатываются прочные условные рефлексы. Однако, существуют и другие центры, раздражение которых вызывает реакцию избегания. Так, крыса с вживленными в эту область электродами после нажима на рычаг избегает находиться рядом с ним. Эти зоны расположены в перевентрикулярных отделах промежуточного и среднего мозга. Области мозга, раздражение которых ведет к подкреплению или избеганию, получили соответственно название центров удовольствия и неудовольствия. Центры неудовольствия найдены в центральном сером веществе, и в гипоталамусе. Структурой мозга, критически связанной со страхом, является миндалина. По мнению П.В. Симонова, вместе с гипоталамусом миндалина образует мотивационно-эмоциональную систему, так как одной из ее функций является выбор доминирующей мотивации. В концепции американского невропатолога Дж. Пейпеца о мозговом субстрате эмоций подчеркивается роль поясной извилины. Изучая эмоциональные расстройства у больных с поражением гиппокампа и поясной извилины, он выдвинул гипотезу о существовании единой системы, объединяющей ряд структур мозга и образующей мозговой субстрат для эмоций. Эта система представляет замкнутую цепь и включает: гипоталамус — передневентральное ядро таламуса — поясную извилину — гиппокамп — мамиллярные ядра гипоталамуса. Она получила название круга Пейпеца. Позднее Мак Лин в 1952 г., учитывая, что поясная извилина как бы окаймляет основание переднего мозга, предложил называть ее и связанные с ней другие структуры мозга лимбической системой. Источником возбуждения в этой системе является гипоталамус. Сигналы от него следуют в средний мозг и нижележащие отделы для инициации вегетативных и моторных эмоциональных реакций. Одновременно нейроны гипоталамуса через коллатерали посылают сигналы в передневентральное ядро в таламусе. По этому пути возбуждение передается к поясной извилине коры больших полушарий. Поясная извилина, по Дж. Пейпецу, является субстратом осознанных эмоциональных переживаний и имеет специальные входы для эмоциональных сигналов, подобно тому, как зрительная кора имеет входы для зрительных сигналов. Далее сигнал из поясной извилины через гиппокамп вновь достигает гипоталамуса в области его мамиллярных тел. Так нервная цепь замыкается. Путь от поясной извилины связывает субъективные переживания, возникающие на уровне коры, с сигналами, выходящими из гипоталамуса для висцерального и моторного выражения эмоций. Однако сегодня гипотеза Дж. Пейпеца приходит в противоречие со многими фактами. Под сомнением оказалась роль гиппокампа и таламуса в возникновении эмоций. У человека стимуляция гиппокампа электрическим током не сопровождается появлением эмоций (страха, гнева и т.п.). Субъективно пациенты испытывают лишь спутанность сознания. П.В. Симонов (1987) относит гиппокамп к "информационному", а не эмоциональному образованию. Из всех структур круга Пейпеца наиболее тесную связь с эмоциональным поведением обнаруживают гипоталамус и поясная извилина. Кроме того, оказалось, что и многие другие структуры мозга, не входящие в состав круга Пейпеца, обладают сильным влиянием на эмоциональное поведение. Среди них особую роль играют лобная и височная кора головного мозга, а также миндалина. Поражение лобных долей приводит к глубоким нарушениям эмоциональной сферы человека. Преимущественно развиваются два синдрома: эмоциональная тупость и растормаживание примитивных эмоций и влечений. При этом в первую очередь страдают высшие эмоции, связанные с деятельностью, социальными отношениями, творчеством. Билатеральное удаление у обезьян височных полюсов ведет к подавлению их агрессивности и страха. Эффект сходен с разрушением миндалины. Передняя лимбическая кора контролирует эмоциональные интонации, выразительность речи у человека и звуковых сигналов у обезьяны. После двустороннего кровоизлияния в этой зоне речь пациента становится эмоционально невыразительной. Согласно современным данным поясная извилина имеет двусторонние связи со многими подкорковыми структурами (перегородкой, верхними буграми четверохолмия, синим пятном и др.), а также с различными областями коры в лобных, теменных и височных долях. Ее связи, по-видимому, более обширны, чем у какого-либо другого отдела мозга. Г. Шеперд (1987) предполагает, что поясная извилина выполняет функцию высшего координатора различных систем мозга, вовлекаемых в организацию эмоций. В составе эмоции принято выделять собственно эмоциональное, субъективное переживание и его соматическое и висцеральное выражение с их самостоятельными механизмами. Диссоциация эмоционального переживания и его выражения в двигательных и вегетативных реакциях обнаружена при некоторых поражениях ствола мозга. Она выступает в так называемых псевдоэффектах: интенсивные мимические и вегетативные реакции, характерные для плача или смеха, могут протекать без соответствующих субъективных ощущений. Таким образом, эмоция — сложное образование, предполагающее интеграцию соматических, вегетативных и субъективных компонентов. Нейронные сети, представляющие эмоции, распределены по многим структурам мозга. О них еще мало что известно. Наиболее полная информация получена о структурах, определяющих знак эмоций. Центры самораздражения, активность которых связывают с появлением положительных эмоций, содержат ДА-ергические нейроны. Эффективность самораздражения соответствует плотности ДА-ергической иннервации. К ДА-ергическим структурам относится медиальный переднемозговой пучок, связывающий гипоталамус и лимбическую систему с лобными долями. ДА-ергические нейроны содержатся также в вентральной тегменталъной области покрышки (ВТО), прилегающем ядре. Через ВТО и прилегающее ядро инициируются и реализуются основные биологические мотивации (нападение, поедание пищи и др.). Черная субстанция и хвостатое ядро (ХЯ) также относятся к ДА-ергической системе. Но они в большей степени обеспечивают реализацию приобретенных форм поведения и причастны к выработке условных рефлексов на положительном подкреплении. В черной субстанции обезьян обнаружены нейроны, реагирующие на пищевое подкрепление соком, а после выработки условного пищевого рефлекса — и на его условные сигналы. Они составили 25% от всех обследованных нейронов этой структуры. Кроме того, эти же нейроны отвечают на новые, неожиданные раздражители. С упрочением условного пищевого рефлекса их реакция на пищевое подкрепление ослабевает, но сохраняется на условный сигнал пищи. Связь активации данной группы нейронов с подкреплением наиболее явно видна при оставлении подкрепления, когда сок подается через 500 мс после реакции животного. Нейроны активировались в момент предъявления пищевого подкрепления, но только после правильно выполненной реакции. В случае неправильных ответов, которые не подкреплялись, отвечали противоположной реакцией — торможением активности в то время, когда обычно давалось подкрепление. Таким образом, на нейронах черной субстанции сходятся активирующие влияния, совпадающие с действием пищевого подкрепления или его условного сигнала, и тормозные влияния, которые соответствуют отмене, отсутствию пищевого подкрепления. В отличие от нейронов префронтальной коры, которые разряжаются в течение всего времени отставления, ДА-ергические нейроны черной субстанции своими фазическими реакциями выделяют основные сигналы, связанные с пищевым подкреплением. В то же время они получают информацию и об отсутствии ожидаемого подкрепления, что проявляется в подавлении их активности. ДА-ергические структуры мозга обладают повышенной чувствительностью к естественным и искусственным опиатам. Это указывает на сложный биохимический механизм центров положительного подкрепления, использующих как медиаторную, так и пептидергическую регуляцию. Так, микроинъекция морфина в ВТО облегчает самостимуляцию мозга, а введение опиатных антагонистов нарушает самовведение героина. Выработанная реакция самостимуляции нарушается и при повреждении ВТО, сопровождающемся снижением уровня ДА. Снижение содержание ДА или повреждение каиновой кислотой его клеток-мишеней для ДА-ергических терминалей нарушает самовведение кокаина. Эффект положительного подкрепления, возникающий при активации обусловлен совместным действием на его нейронах ДА-ергических терминалей от клеток ВТО и опиатного входа. Взаимодействие опиатов с моноаминами на уровне клетки создает качественно новый эффект. Опиоиды открыты и у беспозвоночных. У виноградной улитки ритмическое открывание и закрывание дыхальца определяется совместным действием, ДА и энкефалина. Совместное действие энкефалина и другого медиатора — серотонина — не только обусловливает появление у клетки ритмической активности, но и синхронизацию ее в ансамбле соседних нейронов. Почти все жизненно важные функции организма находятся под контролем опиоидных пептидов. Симонов и его коллеги, исследуя механизм самораздражения латерального гипоталамуса и латеральной преоптической области крысы, выделили два типа нейронов, противоположно связанных с мотивацией и положительными эмоциями. "Мотивационные" нейроны проявляли максимум активности при стимуляции зон самораздражения силой тока, вызывающей мотивационное поведение, и тормозили свою активность со снижением уровня естественной пищевой и водной мотивации. "Подкрепляющие" нейроны достигали максимума активности при стимуляции силой тока, вызывающей самораздражение. Они также возбуждались при естественном насыщении животного. Авторы полагают, что "подкрепляющие" нейроны в центрах самораздражения латерального гипоталамуса и преоптической латеральной области представляют систему "истинной" награды со свойственной ей положительной эмоцией, которая позволяет вырабатывать активные формы условных инструментальных рефлексов, когда животное полностью предотвращает действие болевого раздражителя (рефлекс избегания). П.В. Симонов (1997), рассматривая формирование инструментальных оборонительных рефлексов, различает разные формы награды — в виде ослабления наказания и получения собственно положительного подкрепления, когда активируются центры награды и вовлекается механизм положительных эмоций. Чтобы сформировать инструментальный рефлекс избегания, в результате которого крыса перестает получать наказание током, в качестве подкрепления требуется только награда. Данная форма инструментального рефлекса сочетается с активностью позитивно-подкрепляющих нейронов латерального гипоталамуса, тогда как для выработки классического оборонительного рефлекса или реакции избавления достаточно отмены отрицательного подкрепления. Их осуществление сопровождается усилением активности мотивационных нейронов и подавлением импульсаций — подкрепляющих. При высокой пищевой мотивации активируются нейроны латерального гипоталамуса, а при насыщении — вентромедиального. В латеральном гипоталамусе выявлены глюкозочувствительные нейроны, которые в ответ на электрофоретическое подведение к ним глюкозы пропорционально дозе тормозят свою активность без изменения сопротивления мембраны. В вентромедиальном гипоталамусе найдены глюкозорецептивные нейроны. На глюкозу они реагируют противоположно — в соответствии с величиной дозы увеличивают частоту разрядов. Глюкозочувствительные нейроны латерального гипоталамуса у обезьяны реагируют учащением спайков на вид и запах пищи, на вид шприца с глюкозой, на скорлупу ореха. Их активность модулируется пищевой мотивацией. Только у голодной обезьяны эти нейроны реагируют на пищевые стимулы или их сигналы. Введение глюкозы снижает их активность. По данным Е. Роллса, сначала возникает реакция нейронов, реагирующих на вид пищи (через 15—200 мс), а затем включаются нейроны, отвечающие на прием пищи (300 мс). Обнаружены нейроны, отвечающие либо на вид пищи, либо на вид воды. Глюкозочувствительные нейроны вовлекаются в процесс пищедобывательного поведения. У обезьян их активность резко возрастает за 2—0,8 с перед нажимом на рычаг для получения пищи и подавляется во время движения и следующего за ним подкрепления. Эти нейроны одновременно чувствительны к НА и опиатам. В латеральном гипоталамусе, кроме того, имеются глюкозоне-чувствительные нейроны. Они также активируются во время пищедобывательного поведения, однако их реакции совпадают с движениями, а не предшествуют им. Эти нейроны принимают участие в реализации движений и, кроме того, распознают ключевые экстероцептивные стимулы. Они чувствительны к ДА. Предполагают, что глюкозоне-чувствительные нейроны интегрируют внешнюю информацию, тогда как глюкозочувствительные участвуют в интеграции внутренней информации и подкрепления. Животные, как и человек, имеют тенденцию принимать стимуляторы (кокаин, амфетамин) и опиаты (героин, морфин). Действие этих веществ можно сравнить с действием электрического раздражения на центры положительного подкрепления. Наркотики действуют не только на центры награды, стимулируя их. Параллельно они тормозят центры отрицательного подкрепления (наказания). Поведенчески частично это проявляется в изменении структуры защитных реакций: усиливается роль активно-оборонительного поведения и подавляется пассивная форма защиты, снимаются страх, тревожность. При прекращении введения наркотика возникает отрицательное состояние, которое также стимулирует применение наркотика, чтобы снять это состояние. Состояние эйфории, вызываемое наркотиком, является той положительной эмоцией, которая работает как сильное положительное подкрепление и обеспечивает быстрое формирование устойчивых условных связей данного состояния с различными внешними стимулами, обстановкой и др. Наркотическая эйфория подобна эффекту, вызываемому активацией ДА-ергических центров положительного подкрепления. Ее привлекательность определяется не только ее положительным гедоническим тоном, но и тем, что человек переживает подъем умственной и физической активности, его работоспособность повышается, он мыслит ясно и четко. Необходимым условием для формирования активной формы поведения — инструментального рефлекса — является наличие положительной эмоции в качестве подкрепления. Связь наркотика с положительной эмоцией объясняет механизм инициации поведения "поиска наркотика". Однажды испытав положительный эффект от приема наркотика или электрического раздражения мозга, животное образует ассоциации с сигналами, которые сопутствуют приему наркотика. Так, крысы ищут место, которое ассоциировано с введением наркотика или раздражением положительного центра. Крысы, не имеющие опыта введения опиатов, легко начинают осуществлять его самовведение аналогично тому, как это происходит при самораздражении мозга. Стимулы, сочетаемые с действием морфина, сами могут вызывать ряд реакций, присущих наркотику как безусловному раздражителю. Таким образом, условные раздражители (привычная обстановка) могут привести к активации наркотического состояния и при возвращении к прежней обстановке восстановить потребность в наркотиках. Применение наркотиков и электрическая стимуляция системы положительного подкрепления характеризуются эффектом "разогрева". После подачи электрического стимула возникает стремление усилить самораздражение. А прием однократной дозы наркотика сопровождается увеличением его самовведения у крысы. Эффект самораздражения мозга обнаружен и у улитки. Поведение моллюска изучали при его свободном перемещении по шару, погруженному в воду. При касании улиткой стержня, связанного с электродом, замыкалась цепь подачи тока на определенную группу нервных клеток. Когда стержень не подключен к току, улитка время от времени с некоторой фиксированной частотой касается стержня. В случае если касание стержня замыкает цепь и ток подается на группу клеток оборонительного поведения париетального ганглия, после одного-двух касаний улитка, оставаясь активной, избегает стержня и частота касания падает до нуля. Если ток пропускается через клетки мезоцеребрума, связанные с половым поведением, улитка начинает касаться стержня все чаще и чаще. Одновременно раздражение клеток мезоцеребрума подавляет ответ в клетках оборонительного поведения, что указывает на реципрокные отношения между центром полового "удовольствия" и центром "неудовольствия". Использование в качестве положительного подкрепления прямого раздражения нейронов полового поведения увеличивает вероятность появления подкрепляемой реакции. Так, сочетая спонтанное закрытие дыхальца с раздражением мезоцеребральных клеток, можно увеличить длительность закрытого состояния дыхальца. Сочетание открытия дыхальца с раздражением тех же клеток приводило к увеличению времени пребывания его в открытом состоянии, Раздражение нейронов, запускающих оборонительное поведение, может использоваться в качестве отрицательного подкрепления. 2. Теория Хенри В своей теории эмоций П. Хенри, объясняя происхождение положительных и отрицательных эмоций, выделяет две ортогональные системы активации. Он рассматривает два типа паттернов вегетативных реакций, которые сочетаются с активной или пассивной формами оборонительного поведения, и связывает их оба с функцией миндалины. Электрическая стимуляция ее центрального ядра вызывает реакцию гнева. Поведенчески это проявляется в позе готовности животного к атаке (уши настороженно торчат). На стимуляцию базолатерального ядра миндалины возникает реакция страха: кошка втягивает голову, шипит, уши прижаты. Эмоции гнева соответствует комплекс следующих вегетативных и гуморальных реакций: интенсивное увеличение содержания НА, рост ЧСС и артериального давления. Параллельно увеличивается уровень мужского полового гормона — тестостерона. Уровень кортикостероидов, выделяемых корой надпочечников, существенно не меняется. Комплекс реакций, характерных для страха, иной. Увеличение АД, ЧСС и НА менее выражено. Выброс адреналина заметно преобладает над выбросом НА. Несколько увеличивается уровень кортизола в крови (показателя активности коры надпочечников). В качестве особого типа отрицательных эмоций П. Хенри рассматривает депрессию, которая характеризуется беспомощностью, потерей контроля и отказом от усилий, направленных на преодоление стресса, и общим поведенческим торможением. Причину состояния депрессии он видит в активации системы гипофиз — кора надпочечников. Высвобождаемый гипофизом адренокортикотропный гормон (АКТГ) стимулирует выброс кортикостероидов (в том числе кортизола). Механизм секреции кортизола включает следующие этапы: стрессовый фактор заставляет гипоталамические центры секретировать кортиколиберин, который в свою очередь воздействует на гипофиз, заставляя его вырабатывать кортикотропин. Последний переносится кровью к надпочечникам и стимулирует у них секрецию кортизола. В норме этот процесс прекращается, как только избыток кортизола фиксируется рецепторами глюкокортикоидов в мозге. У больных депрессией кортиколибирин вырабатывается с избытком, так как процесс его подавления нарушен. Для депрессии характерно высвобождение эндорфинов, которые действуют подобно морфину, подавляющему боль. Показатели симпатической активности (АД, НА, адреналин, ЧСС) свидетельствуют о брадикардии. Типы нейроэндокринных реакций, связанных с тремя отрицательными эмоциями, показаны на рисунке ниже.
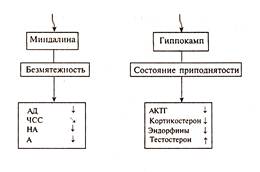
Три типа паттернов вегетативных и гуморальных реакций, связанных с тремя отрицательными эмоциями: гневом, страхом и депрессией. Эмоции различаются уровнем потери контроля над ситуацией. Гнев рассматривается как реакция сопротивления ограничению свободы действия. Страх — реакция на реальную угрозу потерять контроль. Депрессия — состояние беспомощности, поведенческого торможения как следствие сильного дистресса, который не преодолен. Первые две эмоции связаны с функцией миндалины. Депрессия же отражает потерю контролирующей функции системы гиппокамп — септум. Предполагают, что система гипофиз — кора надпочечников находится под контролем гиппокампа, предохраняющего ее от чрезмерной активации, так как после удаления гиппокампа мыши в условиях социальной конкуренции показывают рост кортикостерона. По мнению П. Хенри, для эмоций гнева и страха имеется общий противоположный полюс — положительная эмоция в виде переживания состояния безмятежности, релаксации, которое сочетается со снижением активности мозгового вещества надпочечников (НА, А), падением АД и ЧСС. У животных состояние безмятежности выражается в поведении ухаживания за собой (вылизывании) и другими особями. Противоположность состояния депрессии и беспомощности образует состояние приподнятости с чувством контроля ситуации и ощущением поддержки со стороны социальных партнеров. На вегетативном и эндокринном уровне этому состоянию соответствует снижение уровня АКТГ, кортизола, а также выполняющих защитную функцию эндорфинов. У мужчин растет уровень полового гормона тестостерона, у женщин также вероятен рост половых гормонов эстрогена и прогестерона. Паттерны вегетативных и эндокринных реакций во время состояния безмятежности и подъема настроения показаны на рисунке ниже.
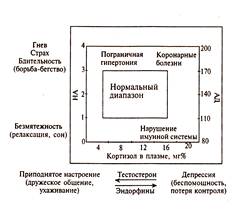
Вегетативные и гуморальные параметры двух эмоциональных состояний: безмятежности — состояния, противоположного гневу и страху, а также состояния приподнятости, противоположного полюсу депрессии. Таким образом, согласно мнению П. Хенри, существуют две биполярные системы: 1) борьба / бегство — безмятежность и 2) депрессия — приподнятое настроение. Все эмоции определяются комбинациями вкладов этих двух независимо функционирующих систем. Существуют многочисленные доказательства независимости активации системы гипоталамус - гипофиз — кора надпочечников от адренергической системы. Вместе с тем это не исключает их параллельное изменение, как у пациентов с депрессией, у которых одновременно с увеличением уровня кортикостероидов в плазме растет содержание НА и А. На рисунке представлена связь эмоциональных переживаний с активацией двух систем: ось катехоламинов (миндалина – симпатическая нервная система — мозговое вещество надпочечников) и ось кортизола (гипофиз — кора надпочечников). За пределами зоны нормальных эмоциональных состояний возникают коронарные и другие нарушения. Высокие значения кортизола стимулируют отклонения в иммунной системе. Увеличение содержания катехоламинов — условие для развития гипертонии. По шкале катехоламинов эмоциональные состояния меняются от высокого уровня бдительности — напряжения в составе эмоции гнева или страха до спокойствия, релаксации. По шкале кортизола эмоциональные переживания меняются от приподнятого настроения (при малых значениях кортизола) до состояния депрессии.
Взаимодействие двух мозговых систем, регулирующих поведение борьбы/бегства и поведенческое торможение. На ординате представлена шкала активации системы: миндалина — симпатическая нервная система — мозговое вещество надпочечников, регулирующая уровень НА и А и оборонительное поведение от борьбы / бегства до релаксации и сна. Она отвечает на активацию синего пятна ростом НА (А) в плазме и увеличением кровяного давления (АД). На абсциссе — шкала активации системы: гипофиз — кора надпочечников, регулирующая уровень кортизола, тестостерона и эндорфинов. Увеличение ее активации приводит к депрессии, для которой характерны высокий уровень кортизола и эндорфина, снижение содержания тестостерона. Агрессия сочетается с высоким содержанием НА и высоким уровнем тестостерона в плазме. Бегство, вызванное угрозой, страхом потерять контроль, связано с высоким содержанием адреналина. Полной потере контроля над ситуацией и поведением соответствует высокий уровень кортизола и эндорфинов и низкий уровень тестостерона, что переживается как депрессия. Нарушения в функционировании холинергической системы (АХ-системы) ведут к психозу с преимущественным поражением интеллектуальных (информационных) процессов. Предполагается, что холинергическая система обеспечивает преимущественно информационные компоненты поведения. Холинолитики — вещества, снижающие уровень активности холинергической системы, ухудшают выполнение пищедобывательного поведения, нарушают совершенство и точность двигательных рефлексов избегания. Уровень серотонина также влияет на эмоции, с ростом его концентрации в мозге настроение у человека поднимается, а его истощение вызывает состояние депрессии. Результаты исследования мозга больных, покончивших с собой в состоянии депрессии, показали, что он обеднен как норадреналином, так и серотонином, причем дефицит норадреналина проявляется депрессией тоски, а недостаток серотонина — депрессией тревоги. Изучение биохимической природы различных эмоций создает далеко не полный портрет эмоциональной сферы. Тем не менее, можно выделить отдельные комплексы медиаторов, гормонов, пептидов и связать их с основными шкалами семантического пространства эмоций, рассматриваемыми в качестве предетекторов эмоционального анализатора. Возможно, что положительный полюс шкалы знаков определяется преобладанием ДА, а другие катехоламины, и прежде всего НА и А, характеризуют отрицательные эмоции. Высоким значениям по шкале ЭА соответствуют высокий уровень как, ДА, так и НА и А. Если катехоламины между собой недостаточно сбалансированы, то шкала эмоциональной активации, вносящая вклад и в положительные и в отрицательные эмоции, замещается одной из частных шкал, более дифференцированно характеризующей эмоции одного знака. В качестве примера можно привести шкалу "радость" — "восторг", представляющую удовольствие, получаемое от игровой ситуации. Можно ожидать, что основу данного эмоционального переживания составляет увеличение выброса ДА, опиатов и АХ. Введение антагонистов мускариновых рецепторов, чувствительных к АХ, и агонистов никотиновых рецепторов подавляет игру. Через агонисты и антагонисты ДА-ергических рецепторов можно манипулировать игрой, так как ДА инициирует игру. Опиаты в низких дозах, воздействующие на ДА-ергические клетки ВТО (вентрально-тегментальной области покрышки), усиливают игровое поведение. Так, введение в ВТО морфина стимулирует моторную активность крыс во время социальной игры, когда они принимают различные позы. Высокие значения шкалы НЭА, совпадающие с переживанием "уверенности", определяются высоким уровнем тестостерона и низким — кортизола и эндорфинов при среднем уровне содержания НА, А и серотонина, а также умеренным количеством, ДА и опиатов. Установлено, что самцы крыс без тестостерона не проявляют должных усилий, направленных на принятие решения. Переживание успеха, достигаемого в деятельности благодаря умениям и компетентности, сопровождается увеличением уровня тестостерона у самцов млекопитающих, включая человека. Имеются данные о том, что у женщин, так же как и у мужчин, тестостерон лежит в основе их эмоциональной устойчивости, которая приходит с достижением профессионального и общественного статуса. В основе депрессии, которую можно связать с состоянием растерянности, т.е. с противоположным полюсом шкалы НЭА, лежит максимальный уровень кортизола и эндорфинов, малое содержание тестостерона и недостаток катехоламинов (НА, ДА и серотонина). Положительный эффект электрошоковой терапии, в 80% случаев устраняющей депрессию у пациентов, связан с усилением синтеза и ростом содержания НА в мозге. Вещества, которые улучшают настроение (ингибиторы МАО), увеличивают содержание НА и ДА в нервных окончаниях. Список используемой литературы 1. Данилова Н.Н. Психофизиология / Н.Н. Данилова. – М.: Аспект Пресс, 1999. – 373 с. 2. Данилова Н.Н. Физиология высшей нервной деятельности / Н.Н. Данилова, А.Л, Крылова. – М.: Учебная литература, 1997. – 432 с. 3. Ильин Е.П. Эмоции и чувства / Е.П. Ильин. – СПб.: Питер, 2001. – 752 с. 4. Смирнов В.М. Физиология сенсорных систем и высшая нервная деятельность / В.М. Смирнов, С.М. Будылина. – М.: Академия, 2003. – 304 с. |